|
Меня всегда интересовал следующий
вопрос: почему у насекомых появилось
полное превращение? Для объяснения
моего недоумения напомню о следующем.
Крылатые насекомые делятся на две
основные группы: насекомые с неполным
превращением и насекомые с полным
превращением.
У насекомого с неполным превращением
из яйца появляется так называемая
"нимфа", то есть маленькая и почти
точная копия взрослого насекомого
(последнее называется "имаго").
Главное отличие нимфы от имаго —
отсутствие органов размножения и
крыльев.
У насекомого же с полным превращением
из яйца появляется личинка, которая
всегда гораздо примитивнее имаго:
например, у мух и у пчёл их личинки
близки по строению к червю, то бишь к
существу с намного более примитивным
устройствам и с намного меньшими
двигательными возможностями, чем
взрослое быстрое крылатое и шестиногое
насекомое.
Мне всегда казалось странным, что
данный вроде бы явный шаг назад — ведь
бо́льшую часть жизни насекомые с
полным превращением стали проводить в
примитивном, в отсталом состоянии —
так вот этот вроде бы явный шаг назад,
во-первых, считается
специалистами не просто шагом вперёд,
но даже "гигантским скачком в
эволюции", а во-вторых,
этот вроде бы явный шаг назад и в
самом деле привёл к несомненно лучшему
демографическому состоянию, чем
демографическое состояние насекомых с
неполным превращением.
В книге Питера Фарба "Насекомые" (этот
американец — почти однофамилец
самого известного в мире энтомолога
француза Жана-Анри Фабра)

про успех и про прогрессивность
насекомых с полным превращением
несколько раз написано примерно
следующее: насекомые с полным
превращением приспособлены к борьбе за
существование лучше насекомых с
неполным превращением потому, мол, что
живут в двух средах.
На мой взгляд, это очень слабое
объяснение успеха полного превращения.
Ибо, во-первых, имеется
немало насекомых с неполным
превращением, которые тоже живут в
двух очень разных средах — например,
подёнки, веснянки, стрекозы —
а во-вторых, к одной
среде, к одним условиям можно
приспособиться намного лучше, намного
полнее, чем к двум и более разнородным
условиям-средам.
Если же я неправ с последним
утверждением, то почему тогда эволюция
насекомых не пошла в предписанном
Питером Фарбом направлении дальше и не
создала виды, которые существуют уже
не в одной и не в двух, а в трёх и
более средах? Почему два — это
наиболее прогрессивное количество сред
обитания одного существа?
В других источниках мне тоже не
удалось найти более-менее
внятное объяснение повышенной
успешности полного превращения —
прежде всего потому, что этот вопрос
просто не ставится или же обходится
стороной (при всём при том что полному
превращению даются самые лестные
оценки). Вот, например, что написано
в Википедии.
"Происхождение полного превращения
Несомненно, полное превращение явилось
гигантским скачком в эволюции
насекомых и всего типа членистоногих.
Относительно его возникновения
существует две основных гипотезы.
Сторонники обеих согласны, что
неполное превращение и,
соответственно, его очевидные
производные (гипо- и гиперморфоз)
произошли от первичного типа
метаморфоза — протоморфоза. Что
касается возникновения
голометаморфоза, то мнения расходятся.
Согласно гипотезе независимого
происхождения, предложенной
М.С.Гиляровым, А.А.Захваткиным и
А.Г.Шаровым, полное превращение
является эволюционным развитием
протоморфоза. При протоморфозе личинки
и имаго обычно живут в почве, под
растительными останками и в других
защищённых местах. Но приобретение
крыльев неизбежно способствовало
переходу имаго к открытому образу
жизни, что, в свою очередь, вынуждало
их нежных личинок тоже жить открыто,
под сильным воздействием
неблагоприятных условий. Возникла
необходимость приспособления личинок к
этим условиям, которое пошло в итоге
двумя путями. В одних случаях развитие
стало всё более проходить под защитой
яйца. Вылупление всё более запаздывало
и происходило на более поздних стадиях
развития, то есть осуществлялась
эмбрионизация развития. Личинки
вылуплялись уже приспособленными к
открытому образу жизни, само же яйцо
стало намного крупнее и богаче
питательным желтком. В других случаях
личинки перешли к ещё более скрытому
образу жизни, благодаря чему у них всё
более усиливались червеобразные
признаки, происходила деэмбрионизация
зародыша. Возник резкий разрыв между
стадией личинки и имаго, который был
восполнен стадией куколки. Так возник
голометаморфоз.
Сторонники второй гипотезы
предполагают, что полное превращение
развилось из неполного. Эти
представления связаны, в первую
очередь, с именами итальянского
учёного А.Берлезе и русского
исследователя И.И.Ёжикова. По их
мнению, личинки насекомых с полным
превращением возникли вследствие
выхода из яйца на более ранних
стадиях развития. Образно говоря,
истинные личинки являются свободно
живущими эмбрионами. Отличие от
предыдущей теории в том, что стадия
куколки не является новой стадией в
полном смысле слова, а соответствует
совокупности нимфальных стадий
насекомых с неполным превращением.
Одной из причин эмбрионизации
считается малое содержание желтка в
яйцах насекомых с полным превращением,
в результате чего личинка не успевает
завершить развитие в яйце и вынуждена
вылупляться раньше. Противоположно,
яйца насекомых с неполным
превращением, как более крупные,
содержат больше питательного желтка.
Поэтому зародыш в своём развитии
достигает более поздних стадий и
вылупляется уже с признаками имаго.
Существует и третья, менее
распространенная гипотеза, согласно
которой истинные личинки являются
прямыми модификациями нимф, то есть
что голометаморфоз развился из
гиперморфоза. Причину такой
модификации видят в дифференцировании
среды́ обитания и образа жизни
личинок и имаго.
Некоторые положения этих гипотез
подкрепляют друг друга, другие
являются взаимоисключающими. Всё это
говорит о недостаточности современных
знаний об эволюции метаморфоза.
Функциональная роль стадий метаморфоза
Для насекомых характерно разделение
функций не только в пространстве
(дифференциация органов), но и во
времени: в течение онтогенеза
происходит не только смена функций
отдельных органов, но меняются и
основные функции организма как целого.
Для личиночных стадий насекомых
характерна функция питания, накопления
пластических и энергетических веществ,
а для имаго — функция размножения.
Функция расселения может выполняться
как имагинальной, так и личиночными
стадиями. Большинству насекомых, как
несомненно прогрессивной группе,
свойственно активное расселение,
осуществляемое полётом. Поэтому весь
облик имаго большей части крылатых
насекомых — это облик расселяющейся
стадии.
Иногда имагинальная стадия выполняет
только функции видовой жизни —
расселения и размножения, утрачивая
функцию питания. Однако выполнение
функций расселения и размножения часто
протекает в различные сроки; часто
расселение происходит в те сроки,
когда половые продукты ещё не созрели.
Так, самки многих совок совершают
перелёты, когда их яичники ещё не
созрели. Более того, после созревания
половых продуктов они отяжелевают
настолько, что теряют способность даже
к небольшим взлётам. Расселительную
функцию иногда имеют и брачные полёты,
особенно у общественных насекомых.
Таким образом, у многих насекомых
можно говорить о возрастном
функциональном разделении стадии имаго
на две "подстадии": расселительную и
репродуктивную.
Утрата имаго расселительной функции
ведёт к регрессивному развитию
взрослых особей. Одним из наиболее
ярких примеров этого являются червецы
и щитовки. Функция расселения у
червецов выполняется личинками
I возраста, так называемыми
бродяжками. Расселение вышедших из яиц
бродяжек совершается в основном
пассивно, переносом ветром, иногда на
очень большие расстояния. Попав в
благоприятные условия на кормовые
растения, подвижные шестиногие личинки
переползают на них и прикрепляются в
наиболее удобных для сосания местах.
Дальнейшее развитие самок
сопровождается впечатляющими
регрессивными изменениями — редукцией,
вплоть до полной утраты, ног, органов
чувств (усиков, глаз), развитием
пассивных органов защиты (щитков) и
прогрессивным развитием репродуктивных
органов (у многих щитовок к концу
яйцекладки тело самки представляет
собой щиток, прикрывающий яйца,
количество которых может исчисляться
тысячами). Утрата расселительный
функции имаго также характерна для
многих паразитических насекомых, к
примеру, для веерокрылых
".
Как можно видеть, тема превосходства
полного превращения над неполным
("Несомненно, полное превращение
явилось гигантским скачком в эволюции
насекомых") в этом тексте
совершенно не раскрыта.
Согласно первой гипотезе, предложенной
М.С.Гиляровым, А.А.Захваткиным и
А.Г.Шаровым, оба вида превращений
имеют независимое друг от друга
происхождение, то есть здесь
принципиально не может быть
последовательной смены, замещения
чего-то менее
прогрессивного чем-то
более прогрессивным.
Согласно второй гипотезе, предложенной
А.Берлезе и И.И.Ёжиковым, полное
превращение развилось из неполного. Но
в чём именно заключается
прогрессивность полного превращения —
совершенно непонятно. Повторяю цитату:
"...личинки насекомых с полным
превращением возникли вследствие
выхода из яйца на более ранних стадиях
развития. Образно говоря, истинные
личинки являются свободно живущими
эмбрионами. Отличие от предыдущей
теории в том, что стадия куколки не
является новой стадией в полном смысле
слова, а соответствует совокупности
нимфальных стадий насекомых с неполным
превращением. Одной из причин
эмбрионизации считается малое
содержание желтка в яйцах насекомых с
полным превращением, в результате чего
личинка не успевает завершить своё
развитие в яйце и вынуждена
вылупляться раньше. Противоположно,
яйца насекомых с неполным
превращением, как более крупные,
содержат больше питательного желтка.
Поэтому зародыш в своём развитии
достигает более поздних стадий и
вылупляется уже с признаками имаго.
"
Здесь объясняется всего лишь
происхождение отличий в развитии
насекомых с полным превращением, но
совершенно не демонстрируется
прогрессивность этих отличий, то бишь
не объясняется, почему появившиеся
отличия в развитии имеют повышенную
выгодность.
И даже, напротив, сама вот эта фраза
"Одной из причин эмбрионизации
считается малое содержание желтка в
яйцах насекомых с полным превращением,
в результате чего личинка не успевает
завершить своё развитие в яйце и
вынуждена вылупляться раньше"
наводит на мысль о как раз ущербности
полного превращения: личинке было бы
лучше "завершить своё развитие в
яйце", но она этого сделать не
может и потому "вынуждена
вылупляться раньше", то есть
раньше, судя по смыслу текста, более
подходящего для вылупления момента.
Третья гипотеза тоже никак не
обосновывает уверения о "гигантском
скачке в эволюции насекомых" — она
всего лишь называет причиной
возникновения полного превращения
дифференцирование "среды́
обитания и образа жизни личинок и
имаго" — а значит, сие то же
самое, на что ссылался Питер Фарб и
что я уже разобрал чуть выше.
Бытует и ещё одно объяснение
повышенной выгодности полного
превращения: мол, при полном
превращении взрослые особи и личинки
потребляют разную пищу и потому не
составляют друг другу пищевой
конкуренции. А внутривидовая
конкуренция — она, дескать, самая
жестокая.
Однако это объяснение является, судя
по всему, не шибко убедительным (и,
видимо, как раз по данной причине не
фигурирует в Википедии): большинство
имаго насекомых с полным превращением
вообще ничего не едят, а в лучшем
случае лишь пьют воду или нектар (как
бабочки), или сок деревьев (как
некоторые жуки).
У многих же иных видов насекомых с
полным превращением имаго питаются
почти в точности тем же самым, что и
личинки (пчёлы, осы, шмели, муравьи,
щитовки, божьи коровки
и т.д.).
А вот у насекомых с неполным
превращением иногда как раз
наблюдается упомянутая разница в
питании нимф и имаго. Например, это
имеет место у стрекоз (водные
насекомые у нимф и воздушные жертвы у
имаго).
Кстати, даже если бабочки ели бы то же
самое, что и гусеницы, то больших
проблем сие, скорее всего, не вызвало
бы. Ведь кормящиеся гусеницы частенько
образуют настолько большие и густые
скопления, что совершенно очевидно: у
этих многочисленных существ в еде нет
никакого недостатка.
Итак, вот оно, моё совершенно
очевидное, лежащее на поверхности
объяснение прогрессивности иногда
очень и очень значительной
примитивизации доимагинальных стадий у
насекомых с полным превращением (то
есть у насекомых с червеобразными
личинками типа бабочек, брачнокрылых и
мух).
Всё дело, действительно, в жизни
личинки и взрослой формы в разных
средах. Но только я имею в виду уже не
абстрактно разные сре́ды, то есть
не как попало разные сре́ды, а
такие разные сре́ды, каждая из
которых очень особенно, очень точно
подходит для главных функций у разных
стадий развития насекомого.
Личиночная стадия — это, как
совершенно правильно отмечено в тексте
из Википедии, стадия "накопления
пластических и энергетических
веществ", а взрослая стадия — это
стадия размножения и, как правило,
расселения.
Для масштабного и в то же время
быстрого расселения потомства по
планете лучше всего подходит почти
пустая воздушная среда. Эта же
воздушная среда лучше всего подходит и
для максимально быстрого нахождения
дальнородственного полового партнёра
(сия дальнородственность полового
партнёра позволяет резко усилить
полезную изменчивость и в то же время
избежать опасностей инбридинга).
А вот для "накопления пластических
и энергетических веществ"
воздушная среда подходит плохо: в ней
относительно мало питательных веществ,
но в то же время в ней хороший доступ
к добыче для всевозможных хищников.
Разумеется, не только тех хищников,
что летают, но и тех, которые, как
лягушки или как
рыбы-брызгуны и как
молодые арапаймы, внимательно следят с
земли или из-под воды за
всеми передвижениями по воздуху
потенциальной добычи.
В то же самое время существуют такие
сре́ды, в коих быстрое и далёкое
передвижение (необходимое, напоминаю,
для расселения потомства или для
размножения путём скрещивания именно с
дальнородственными половыми
партнёрами) сильно затруднено, но
которые зато позволяют хорошо
скрываться от хищников и в то же время
сами полны "пластических и
энергетических веществ".
Я имею в виду не только верхние слои
почвы с их огромной массой живых и
разлагающихся растений, но и сами
ветки растений, по коим ползают хорошо
замаскированные гусеницы.
Кроме того, я имею в виду трупы
животных или даже живые организмы, на
которых или внутрь которых откладывают
яйца наездники, одиночные осы и оводы.
Эта богатая питательными веществами
среда не только не требует быстрых
передвижений, но и не допускает их
вследствие своей большой вязкости.
Соответственно, такие органы
передвижения, как относительно тонкие
и длинные, быстрые и
высокоманёвренные ноги, оказываются,
во-первых, бесполезными
(среда почти не допускает их
использования), а
во-вторых, излишними
(материалы для строительства этих ног
будут использованы впустую).
У насекомых же с неполным превращением
такой выгодной специализации, такой
полезной для успешного выживания
способности оказываться в нужное время
в нужных средах обычно нет.
Соответственно, неполное превращение
вынуждает доимагинальные формы жить,
как правило, почти в той же самой
среде, что и имагинальные формы. А это
для ещё слишком слабых, слишком
неподготовленных к полноценной борьбе
за выживание доимагинальных форм
слишком часто оказывается неподъёмной
задачей.
Однако написанное выше — это далеко не
весь мой ответ. Ведь я знаю, что
остался ещё один довольно хитрый
вопрос: а как обстоят дела
с прогрессивностью-регрессивностью
полного-неполного
превращения насекомых в том случае,
если эти насекомые — общественные? То
бишь можно ли найти прогрессивные
черты у муравьёв-ос-пчёл,
которые проходят полное превращение,
если сравнивать их с термитами, у
которых превращение неполное?
Похоже, что таких очевидно
прогрессивных черт — нет. И даже,
напротив, у термитов имеется то
преимущество, что их нимфы могут
намного раньше включаться в общую
трудовую деятельность термитника.
В то время как у муравьёв, у шмелей и
у общественных ос и пчёл (а то ведь у
двух последних большинство видов — не
общественные) их личинки почти не
включаются в общую трудовую
деятельность до момента выхода из
куколок (только у некоторых муравьёв
их личинки используются как "швейные
машинки" для склеивания выделяемыми
ими, личинками, шёлковыми нитями краёв
листьев, получающиеся "кулёчки" из
которых становятся жилищами данного
вида муравьёв). Они лишь смирно лежат
в специально отведённых местах и
требуют постоянного ухода типа подноса
корма, уборки экскрементов и
вылизывания.
Правда, общественные брачнокрылые
заселили почти всю земную сушу — кроме
зоны вечных льдов и мерзлоты, — в то
время как термиты являются в основном
экваториальными-субэкваториальными
и лишь в очень немногих случаях
субтропическими насекомыми.
Но данное обстоятельство, похоже, уже
никак не связано с
преимуществами-недостатками
полного и неполного превращений.
Вадим Протасенко:
А меня всегда интересовал другой
вопрос: как эволюция вообще смогла
породить насекомых с полным
превращением? Казалось бы, никаким
постепенным, никаким поэтапным
накоплением полезных мутаций сие
сделать нельзя (впрочем, у меня просто
не было времени подумать об этом).
Ведь все многоклеточные животные
проходят развитие от зародыша через
множество примитивных форм, то есть
каждый раз как бы проживают весь путь
эволюции — но только внутри яйца или
утробы. Просто у насекомых с полным
превращением зародыш на
какой-то стадии покидает
яйцо и живёт вне его, пока не
превращается в половозрелое насекомое.
Но, с другой стороны, в яйце или в
утробе идёт поступательное развитие
плода через все исторические формы
животного (в том числе и у
земноводных, которые также проходят
промежуточную стадию вне яйца), а
насекомое с полным превращением в
стадии куколки разбирает себя до
органических молекул и потом собирает
уже в половозрелое насекомое. Вот этот
процесс у меня и не укладывается в
голове.
Каким образом, повторяю, такое могло
возникнуть в ходе эволюции как череды
последовательных мелких изменений?
Материалист:
Уважаемый Вадим, насколько мне
известно, на самом деле в стадии
куколки не происходит совсем уж
полной, "до органических молекул"
"разборки" личинки — например, почти
неизменной остаётся её нервная система
(благодаря чему бабочка помнит то,
чему обучилась гусеница), а кроме
того, сохраняют клеточную целостность
так называемые "имагинальные зачатки",
то есть группы клеток, из которых
формируются органы имаго.
Кстати, если взглянуть на куколок ос
или пчёл и на куколок жуков — причём
находящихся ещё на самых первых
мгновениях сброса личиночных покровов,
— то видно, что покровы этих куколок
уже почти полностью повторяют формы
будущих имаго. Повторяют, ещё раз
обращаю внимание, именно сразу, именно
с ходу, поскольку формируются
буквально за несколько часов, без
многомесячного ожидания полного
преображения во взрослое насекомое.
Ничего особенного, ничего совсем уж
удивительного в разборке, в
"растворении" личиночных тканей тоже
нет.
Ведь в живом мире повсеместно
распространено такое явление, как
апоптоз, то есть умирание, точнее,
растворение одной клетки по "приказу"
(издаваемому, естественно, в
химической форме) окружающих её
соседок.
У насекомых с полным превращением этот
самый апоптоз претерпел просто
количественные изменения.
Ещё раз: апоптоз у насекомых с полным
превращением — он отличается от
апоптоза насекомых с неполным
превращением именно чисто
количественно. То есть у насекомых с
неполным превращением апоптоз
(растворение структурных элементов
организма до молекул) и дальнейшее
строительство из них новых структур —
он тоже есть. Но только меньший по
масштабу.
И вообще, деление насекомых по полноте
превращения — оно в целом довольно
условно. Например, стрекозу
систематики относят к насекомым с
неполным превращением — хотя её
личинка по многим параметрам (аппарат
питания, аппарат движения, аппарат
дыхания, устройство кишечника,
двигательные навыки) совершенно не
похожа на имаго.
А вот ручейника систематики относят к
насекомым с полным превращением, хотя
его куколка — равно как и куколка
комара — всё видит и неплохо плавает,
двигая ногами, как
клоп-гладыш.
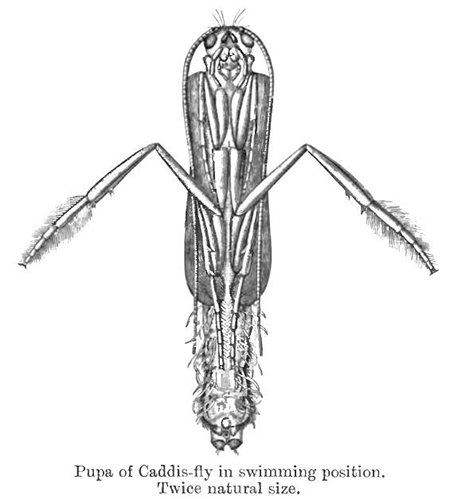
То есть у этих куколок не растворяются
не только нервная система и
имагинальные зачатки, но и глаза, и
мышцы ног, и мышцы брюшка.
Почему же тогда не отнести к насекомым
с полным превращением и стрекозу:
условившись, что её личинка на
последней стадии перед вылетом — это
тоже плавающая куколка?
Так что полное превращение совершенно
нормально возникло путём постепенного
накопления мелких полезных мутаций.
Возможность этого полного превращения
как стадии в развитии со значительным
демонтажом одних структур и монтажом
из получившегося вещества других
структур возникла благодаря
самому́ устройству членистоногих.
У которых опорные структуры ("скелет")
— внешние и не растущие.
Соответственно, у всех членистоногих
изначально имелась потребность в
линьке, то есть в сбросе старых и
обычно отстающих в размерах покровов.
Линька же требует, понятно, длительной
остановки обычной жизнедеятельности:
поскольку покровы животному нужны чаще
всего именно жёсткие. Но для вылезания
из старых покровов и для последующего
увеличения в размерах новые, уже
имеющиеся на теле покровы должны быть
ещё мягкими.
Вот за время этой остановки обычной
жизнедеятельности и происходит
необходимое затвердение новых
покровов.
И если неблагоприятные обстоятельства
нарушат покой и тем самым помешают
правильному по форме затвердению, то
новые покровы, новый экзоскелет может
оказаться уродливым, не позволяющим
полноценно жить дальше.
Именно из этой вынужденно долгой
остановки во время любой линьки,
несомненно, и берёт своё начало полное
превращение как не обычная маленькая
перестройка организма — типа той, что
имеет место у тараканов, а почти
тотальная — типа той, что имеет место
у мух.
Кстати, повторюсь, даже у насекомых с
неполным превращением последняя линька
тоже приводит к принципиальным, к
качественно крупным (возникновение
половой и летательной систем)
изменениям организма. Хотя "зародыши"
этих изменений (типа появления
зачатков крыльев) формируются ещё в
предпоследней линьке.
Так что полное превращение было
сформировано эволюцией путём всё же
совершенно нормального и вполне
постепенного, последовательного,
поэтапного, поштучного собирания в
кучку всех необходимых для
половозрелой формы изменений тела и
поведения именно в рамках самой
последней линьки.
17.12.2013 г.
|